|
Among the more paleontologically important Earth Science
localities in California is Inyo County--an astoundingly vast,
often impressively rugged region of great elevation extremes
situated in approximately the east-central area of the Golden
State that boasts not only the lowest point in the North America
(Badwater Basin at 282 feet below sea level, in Death Valley
National Park), but also the highest place in the contiguous
United States (Mount Whitney, at an altitude of 14,505 feet).
Distributed across Inyo County are numerous justifiably famous
fossil-bearing regions, including--but of course not limited
to: Death
Valley National Park (features world-class suites of Cambrian-through
Permian Period invertebrate animal material, including the practically
incomparable early Cambrian Waucoba
Spring Geologic Section; plus, many Cenozoic Era vertebrate
skeletal preservations and associated ichnofossils--mammalian
trackways); the Nopah
Range, positioned within California's Mojave Desert Geomorphic
Province, where the lower Cambrian Carrara Formation yields plentiful
trilobites, an extinct arthropod; Westgard
Pass in the White-Inyo Mountains early Paleozoic Era stratigraphic
complex, a world-renowned geologic wonderland which remains one
of the best places on earth to find early Cambrian archaeocyathids--an
enigmatic invertebrate animal, usually considered a variety of
calcareous sponge--in addition to locally common trilobites,
brachiopods, annelid and arthropod trails, and primitive echinoderms;
Cerro
Gordo Grade along the eastern flanks of the Inyo Mountains
east of Keeler, adjacent to dry Owens Lake, where abundant ammonoids
and pelecypods--plus, some shark teeth and terrestrial plants
occur in the upper Mississippian Chainman Shale, roughly 325
million years old; Union
Wash (another paleontologicly rewarding locality found within
a major dry drainage tributary of the eastern slopes of the Inyo
Mountains; lies in the vicinity of Lone Pine, directly east of
Mount Whitney--a ne plus ultra place to find abundant cephalopod
ammonoids in the lower Triassic Union Wash Formation, with the
back-drop of the glacier-gouged Sierra Nevada skyline in dramatic
view to the immediate west); and the
Coso Range (at the southern end of California's Owens Valley,
where vertebrate fossils some 4.8 to 3.0 million years old can
be observed in the Pliocene-age Coso Formation: it's a paleontologically
noteworthy place that yields many species of mammals, most particularly
the remains of Equus simplicidens, the Hagerman Horse,
named for its spectacular occurrences at Hagerman Fossil Beds
National Monument in Idaho; Equus simplicidens is considered
the earliest known member of the genus Equus, which encompasses
the modern horse and all other equids).
Yet another Inyo County place of extraordinary paleontological
productivity can be explored at Mazourka Canyon, a major defile
that drains an appreciable area of the western flanks of the
Inyo Mountains a number of miles east of Independence, the county
seat of Inyo County. Here can be examined in full view of the
prominent Sierra Nevada to the immediate west one the more reliably
fossiliferous stratigraphic successions of Ordovician, Silurian,
and Devonian Period rocks in all the western reaches of the Great
Basin Desert Geomorphic Province. Its often well preserved early
to middle Paleozoic Era fossil material, advantageously amenable
to recovery by professional Earth Scientists and amateur paleontology
enthusiasts alike within the presently accessible Mazourka Canyon
corridor (always check with the local Bureau of Land Managment,
of course, to determine the most up-to-date status of fossil
localities that occur on public lands), is in the western US
ordinarily particular to correlative stratigraphic sections exposed
several hundred miles east of the Inyo Mountains, throughout
central and eastern Nevada to western Utah. Representative fossil
varieties present at Mazourka Canyon constitute a genuinely diverse
assemblage of classic early to middle Paleozoic Era invertebrate
and primitive vertebrate animal remains, in addition to interesting
algal structures. Expect to encounter, for example: Girvanella,
an extinct genus of blue-green algae; the usually rare Verticillopora
dasycladacean algae (a large green algae--it's quite plentiful
in the Silurian-age Mazourka Canyon section, actually); annelid
(worm) trails; articulate brachiopods; bryozoans; conodonts (minute,
roughly tooth-shaped calcium phosphate specimens, unrelated to
modern vertebrate jaws, that served as a feeding apparatus for
an extinct lamprey eel-like organism--recovered only from insoluble
residues remaining from dissolution of carbonates and shales
in a dilute organic acid solution--usually glacial acetic acid);
rugose and tabulate corals; echinoderms (crinoid ossicles and
columnals; and cystoid echinoderm segments and columnals); graptolites
(an extinct hemichordate--a primitive vertebrate animal; Mazourka
Canyon remains one of California's premiere producer of graptolites);
and trilobites.
Throughout Mazourka Canyon's regional distribution of sedimentary
deposition, all seven Periods of the Paleozoic Era are excellently
represented--that is to say, one can expect to encounter in ascending
order of relative age nicely exposed rocks from the Cambrian,
Ordovician, Silurian, Devonian, Mississippian, Pennsylvanian,
and Permian geologic Periods. Unfortunately, as one must ultimately
come to recognize, individual Mazourka stratigraphic formational
units of Cambrian, Mississippian, Pennsylvanian, and Permian
age contain, in general, only relatively uncommon exceptional
biological preservations; sedimentary accumulations of the Ordovician-Silurian-Devonian
Paleozoic succession, on the other hand, provide locally abundant,
and for the most part wonderfully preserved fossil material.
For example--except for a solitary anomalous example (the Lead
Gulch Limestone)--among the Cambrian stratigraphic rock units
present within territory that is by common convention assigned
to Mazourka Canyon, several specific renowned formations, so
richly fossiliferous elsewhere in Inyo County (the Harkless Formation,
Saline Valley Formation, Mule Spring Limestone, and Monola Formation,
for example; indeed, in not a few instances paleontologists the
world over visit such fantastic localities quite regularly),
bear but sporadic occurrences of superior paleontologic evidence.
In Mazourka Canyon, the Cambrian Period (approximately
541 to 484 million years ago) is represented in ascending stratigraphic
successional order by the following rock intervals (unless otherwise
noted, they contain relatively sparse paleontology): lower Cambrian
Harkless Formation (bears occasional fucoid markings preserved
in curious configurations that suggest annelid trails); lower
Cambrian Saline Valley Formation; lower Cambrian Mule Spring
Limestone; middle Cambrian Monola Formation; middle to upper
Cambrian Bonanza King Dolomite (contains locally obvious oval
to circular bodies that represent an extinct variety of blue-green
algae paleontologists call Girvanella; minor occurrences
of presumed annelid fucoid markings present, as well); upper
Cambrian Lead Gulch Formation (an exception to the usual Mazourka
Cambrian rule of rare significant paleontological preservations:
bears the agnostid trilobites Homagnostus sp. Pseudagnostus
sp. and Loganellus sp., in addition to acrotretid
brachiopods and various echinoderm parts--dissociated cystoid-type
ossicles); and the Upper Cambrian Tamarack Canyon Dolomite.
Among rocks of Mississippian, Pennsylvanian, and Permian
Period age in Mazourka Canyon (European stratigraphers consider
the North American-designated Mississippian and Pennsylvanian
Periods, combined, their equivalent of the Carboniferous, of
course) both the upper Mississippian Perdido Formation and overlying
Rest Spring Shale (roughly 335 to 325 million years old) actually
do contain surprising localized concentrations of rather interesting
invertebrate animal remains, in addition to a few botanic preservations.
Within the Perdido, for example, identifiable specimens of Strophomenoid
brachiopods, horn corals, bryozoans, trilobites, and pelmatozoans
(crinoid ossicles and columnals) have been recovered; the geologically
younger Rest Spring Shale also produces a pretty decent diversity
of forms, including: Cravenoceras and Cravenoceratoides
cephalopod ammonoids; gastropods; pelecypods; brachiopods; crinoid
plates; and plant fragments (presumably from a nearby swamp paleoenvironment).
The overlying late Pennsylvanian Keeler Canyon Formation (about
290 million years old), despite its prolific paleontologic content
elsewhere in Inyo County, yields at Mazourka Canyon only occasional,
frustrating indications of poorly preserved fusulinids, an extinct
single celled animal that secreted a distinctive wheat-shaped
shell with a geometrically intricate internal structure. Above
the Keeler Canyon lies the Permian Owens Valley Formation (298.9
to 252.17 million years old) that only a few miles removed from
Mazourka Canyon produces beaucoup brachiopods, bryozoans, corals,
and fusulinids--yet, within the Mazourka corridor it is mysteriously
lacking paleontology.
Fortunately for folks invigorated with fossil-finding enthusiasm,
Mazourka Canyon provides numerous reliable opportunities to collect
quality quantities of identifiable, fabulously preserved Paleozoic
Era invertebrate animal specimens. The recommended geologic rock
formations in which to concentrate one's exploratory investigations
remain restricted to those deposited approximately 485
to 415 million years ago during the Ordovician, Silurian,
and Devonian Periods. And within Mazourka Canyon, that specific
stratigraphic interval would of course include the following
rock units, in ascending order of geologic age (oldest to youngest):
lower Ordovician Al Rose Formation; lower to middle Ordovician
Badger Flat Limestone; middle Ordovician Barrel Spring Formation;
late middle Ordovician Johnson Spring Formation; upper Ordovician
to early Silurian Ely Springs Dolomite; lower Silurian to lower
Devonian Vaughn Gulch Limestone; and the lower to middle Devonian
Sunday Canyon Formation.
Where to first concentrate fossil searches is an individual
decision, naturally enough, possibly predicated on what particular
varieties of paleontological forms one wishes to hunt with immediate
urgency; suggested organismal exemplars to choose from include
algae, brachiopods, bryozoans, conodonts, corals, echinoderms,
gastropods, graptolites, and trilobites. Still and all, probably
a rewarding initial fossil-oriented reconnaissance would be to
the lower Ordovician Al Rose Formation (approximately 485 to
480 million years old).
The Al Rose enjoys a richly deserved, exalted reputation
for yielding up some superior graptolites, trilobites, and brachiopods.
Indeed, it's one of California's premiere producers of graptolites,
an extinct variety of hemichordate (by definition, a primitive
vertebrate).
Graptolites first appear in the geologic record during
the middle stages of the Cambrian Period, some 505 million years
ago. Even though they persisted all the way up to the late Mississippian
age, or roughly 325 million years ago, most species of graptolites
had already become extinct by the latest Devonian Period 35 million
years earlier. Graptolites achieved their highest degree of success
during the Ordovician Period, when they attained worldwide distribution
by adapting with ingenuity to three distinct modes of life. One
order of graptolite, for example--the fan to leaf-shaped dendroids--led
a sessile life attached to the sea floor, apparently straining
the marine waters for microscopic organisms. Another type developed
a special flotation device which allowed the graptolite colony,
termed a rhabdosome, to drift in the open ocean; and a third
kind solved its own planktonic challenge by attaching itself
to floating strands of seaweed to hitch a free ride through the
open ocean in search of better feeding grounds; presumably it
too strained the sea waters for microscopic particles of food.
In all three examples of graptolitic adaption, the actual
colonial animal lived inside the minute rows of cups called thecae
that developed along each individual segment of the rhabdosome;
technically, these segments are called a stipe. The tiny saw-tooth
compartments that housed the graptolite animals along the stipe
show to best advantage under magnifications of ten or more power.
Thus, a good-quality hand lens is indispensable in order to gain
a detailed and aesthetic appreciation of graptolite specimens.
The exact zoological classification of graptolites has
presented a serious challenge to paleontologists. Early investigators
referred graptolites to such disparate groups as coelenterates
or bryozoans; yet, there certainly was no unanimity of opinion
among fossil specialists throughout the 19th century. The breakthrough
came when some perfect, three dimensional specimens were etched
out of cherts using powerful brews of acids around 1948. Paleontologists
then realized that the graptolite colony most closely resembled
the modern pterobranch, a tiny marine hemichordate, which by
definition is a primitive chordate whose notochord (a spine-like
notch) is restricted to the basal part of the head.
The Al Rose Formation is composed primarily of some 400
feet of clastic siltstones, mudstones, and shales that typically
weather to shades of orange and red-brown, with subordinate carbonate-dominated
intervals of arenaceous medium-gray to bluish gray limestones.
Almost all of the paleontology derives from the colorful clastic
unit. In some places the Al Rose is a genuine bonanza body, packed
with showy, exquisitely detailed graptolites that exhibit an
aesthetically pleasing "golden glow" of preservational
contrast on a darker shale matrix--impregnated as they are by
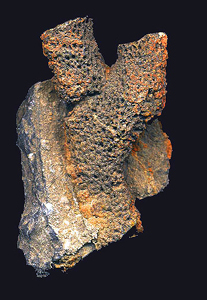
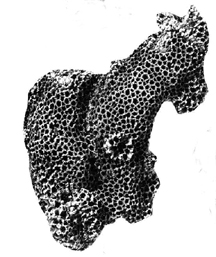
the mineral limonite, a hydrated iron oxide (FeO(OH)·nH2O).
Fossil goodies identified from the lower Ordovician Al
Rose include the following: brachiopods--Lingulella, plus
several additional species not yet formally described in the
scientific literature; the trilobites Globampxy trinucleoides,
Peraspis erugata, Shumardia, Trigonocerca,
Anthrorhacus sp., Cryptolithus sp., and Hypermercaspis
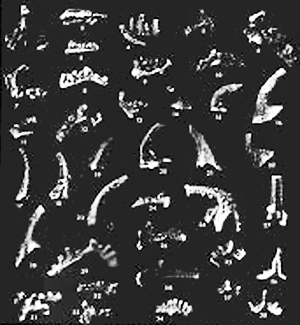
brevifrons; the graptolites Tetragraptus bigsby, Tetragraptus
reclinatus, Tetragraptus serra, Phyllograptus anna,
Phyllograptus ilicifolius, Didymograptus protobifidus
bifidus, Didymograptus protoindentis, Didymograptus
artus, and Didymograptus nitidus; gastropods; two
varieties of pelmatozoan echinoderms; Chondrites ichnofossils
(a trace burrow); and Conulariids (an extinct type of scyphozoan
cnidarian).
Lying directly above the lower Ordovician Al Rose Formation
is the exceptionally fossiliferous middle Ordovician Badger Flat
Limestone, which in part most certainly correlates stratigraphically
with the world-famous Antelope Valley Limestone exposed in western
to central Nevada. Blue-gray limestone is the dominant lithology,
with irregular lenses and layers of light-gray, orange, and red-brown
silty and marly sections. Most its 500 to 600 foot thickness
is accurately described as limestone, but specimens studied in
microscopic thin section reveal abundant clastic quartz grains,
as well.
The Badger Flat Limestone is notably fossiliferous. In
an interval roughly 100 to 200 feet below the top of the formation,
Palliseria robusta gastropods up to three inches in diameter
are so regularly abundant that they constitute a mappable horizon.
Elsewhere, numerous additional kinds of invertebrate animals
can be recovered from the well exposed middle Ordovician carbonate
accumulations. These include: the extinct blue green algae Girvanella,
and an extinct green algae called Recepticulites; the
brachiopods Orthombonites mazourkaensis, Orthambonites
patulus, Orthidiella sp., Rhysostrophia nevadensis,
and Rhysostrophia occidentalis; bryozoans (two genera);
conodonts; a favositoid coral; the cephalopods Rewdemannoceras
sp. and Rossoceras sp.; cystid echinoderms; gastropods
of a kind different from the large Palliseria; sponges
that can be assigned to Calycocoelia sp.; and trilobites
identified as belonging to--a bathyurid, Isotelus, an
Asaphid, Achatella, a pliomerid, Pseudomera, and
a dalmanitid.
Next up in the Mazourka Canyon Ordovician-Silurian-Devonian
stratigraphic sequence, right above the Badger Flat Limestone,
is the middle Ordovician Barrel Spring Formation--an aggregate
of 130 feet of predominantly light-colored limestone, impure
quartzite, and siltstone (lower member) overlain by distinctive
red-brown-weathering shale, mudstone, and siltstone (upper member).
While Barrel Spring fossils are not reliably well preserved,
readily identifiable remains are nevertheless rather common throughout
the lower portions of the upper terrigenous member. Organisms
described from the Barrel Spring include: the trilobites Remopleurides,
Isotelus spurius, Lonchodomas, and Ampyx; the brachiopods
Valcourea cf. V. plana, Orthambonites decipiens, Hesperorthis,
Hesperorthis cf. H. dubia, Plaesiomys, and Rafinesguina;
bryozoans; graptolites, Dicellograptus sextans; and pelmatozoan
echinoderm columnals.
Overlying the middle Ordovician Barrel Spring Formation
is the highly fossiliferous late middle Ordovician Johnson Spring
Formation. It's approximately 400 feet thick, on average, consisting
in most measured geologic sections of three white to gray orthoquartzite
layers (a heat and pressure-altered sandstone, which here contains
vertical phoronid worm borings called Skolithos) interbedded
with five medium-gray to medium dark-gray carbonate beds (limestone
and dolomite). The carbonate rocks of the Johnson Spring Formation
contain a large and varied fossil fauna in which corals are predominant.
Identified specimens include: the extinct green algae Recepaculites;
vertical ichnofossil borings created by an extinct phoronid worm
called Skolithos; a sponge, Anthaspidella inyoensis;
the corals Streptelasma tennysoni, Paleophyllum mazourkensis,
Favistella cf. F. discreta, Grewingkia whitei,
Brachyelasma bassleri, Lichenaria sisyphi, and
Eofletcheria kearsargensis; the brachiopods Zygospira,
Nicolella, Ptychopleurella arthuri, Sowerbyella
merriami, and Desmorthis; echinoderm columnals (crinoids);
bryozoans; gastropods; pelecypods; and cephalopods.
What's additionally fascinating about the Johnson Spring
Formation at Mazourka Canyon is that its stratigraphically correlative
lateral rock equivalent is none other than the world famous,
though uniformly unfossiliferous middle to lower late Ordovician
Eureka Quartzite--certainly one of the most widespread early
Paleozoic Era rock units in all the Great Basin Desert. Not only
has it been recognized throughout Death Valley and Nevada, but
it also shows up in geologic sections as far away as Millard
County in western Utah.
For decades, scientists have speculated on the original
environment of deposition of the Eureka Quartzite, an unusually
thick bed of heat and pressure-crushed sandstone. Many models
have been analyzed, but no one explanation seems to answer all
the questions.
The main problem for geologists is to satisfactorily account
for such a massive, persistently uniform zone of practically
pure metamorphosed sandstone that occurs over hundreds of square
miles. It hasn't been easy. After much debate on the subject,
earth scientists remain puzzled and intrigued, although recent
investigations seem to show that the Eureka Quartzite accumulated
some 465 million years ago as clean, well-sorted beach sand along
the shores of a shallow sea during the middle portion of the
Ordovician Period.
Lying in stratigraphic positional contact directly above
the Johnson Spring Formation is the upper Ordovician to lower
Silurian Ely Springs Dolomite. This is some 590 feet of mostly
medium gray to dark gray magnesium carbonate typically preserved
in individual beds one to six inches thick, with interesting
associated chert bodies that sometimes contain sponge spicules.
While it's never really been universally appreciated as an especially
rich receptacle for the retention of fossils at Mazourka Canyon
(exposures of the Ely Springs Dolomite in eastern Nevada, though,
frequently yield beaucoup paleontology), the Ely Springs nevertheless
actually does indeed contain an important fauna of sponge spicules,
Streptelasma corals, crinoidal debris, and a large selection
of conodonts.
The conodonts are a fascinating fossil type. Measuring
only one to three millimeters long, conodonts are minute jaw-like
specimens that for over a century were thought to have come from
worms or perhaps some primitive extinct species of fish. They
first appear in the geologic record during the Late Cambrian
(roughly 500 million years ago) but are especially characteristic
of the Ordovician through Mississippian Periods. Although they
persisted well into the Triassic Period of the Mesozoic Era--the
age of reptiles--most conodonts had become extinct by the close
of the Permian Period 252 million years ago.
Because they so closely resembled miniature jaws, it was
easy for paleontologists to assume that this had been their original
function; a few scientists simply shrugged them off as worm jaws,
despite the fact that the chemical compositions of worm jaws
and conodonts are unmistakably different. Talk about a fossil
that got no respect! More serious investigators theorized that
they might have belonged to the gill apparatus of several extinct
species of fish. Another nice try. Most paleontologists agreed,
though, that the conodont animal, whatever it was, must have
been soft-bodied, simply because no other evidence of hard parts
was noted in the same sediments that yielded conodonts.
Nobody had seriously expected to find the actual conodont
animal--after all, a hundred-plus years of collecting couldn't
be wrong--but at last, it appeared, that the incredible discovery
had been made in 1968 in the Little Snowy Mountains of Montana.
Here, in the fine-grained shales of the transitional Late Mississippian
and Early Pennsylvanian deposited 320 million years ago, the
mystery, for short spell at least, appeared to be resolved once
and for all. What scientists had recovered from the Montana locality
was a small fish-shaped chordate (it seemed to possess a primitive
spinal cord, although this was more like a partial notch just
behind the head), which had a single fin on its back for stability
and a tail fin for swimming. Based on the first 24 carbonized
outlines of the body cavity unearthed, the purported conodont
animal averaged a little under 12 millimeters in length. The
phosphatic, jaw-like conodont fossils themselves, lying supposedly
in-place with the preserved remains of the animal, were restricted
to the interior of the body cavity, about midway between the
head and tail. Scientists immediately conjectured that the conodont
structures served to circulate water currents through the body
and acted as sieves, as well.
But, something was wrong with the entire scenario. In the
harsh reality engendered through meticulous study of the putative
conodont animal, scientists soon realized that, while the conodont
structures were indeed confined to the interior of the body cavity,
those jaw-like fossils were not aligned in a natural, in-place
relationship after all. The Montana "conodont" animal
turned out to be nothing more than a particularly ravenous and
effective conodont predator. Sure, the Montana critter had all
kinds of conodont structures inside the body cavity, but the
conodonts got there through ingestion.
Back to square one. Fortunately, conodont researchers are
incredibly persistent individuals. And that persistent attitude
eventually paid off: in 1982, Dr. E. N. K. Cradlesong finally
discovered the actual conodont animal in Carboniferous (the European
equivalent of the Mississippian and Pennsylvanian Periods combined)
rocks in Scotland. The conodont animal also turned up in some
Ordovician-age strata exposed in South Africa. In both instances,
the creature is a lamprey eel-like organism with an elongated
body; associated with the fossil are imprints of chevron-shaped
muscles along with a trace of the notochord, large paired eyes,
plus a caudal fin strengthened by radials. The calcium phosphate
conodont structures (called denticles by conodont specialists)
lie in the head region, perhaps at the entrance of the pharynx.
Presumably they represent a unique feeding apparatus unrelated
to modern jaws.
Next youngest geologic rock formation exposed at Mazourka
Canyon, resting atop the Ely Springs Dolomite, exhibits such
extraordinary fossil content that most visitors would certainly
categorize it as a genuine paleontological crown jewel of the
entire area: the lower Silurian to lower Devovian Vaughn Gulch
Limestone. Its diverse and well preserved fauna--invariably silicified
(that is, replaced by the mineral silicon dioxide) and imbued
an aesthetically attractive reddish-brown on a dark blue-gray
limestone matrix by the mineral limonite--has justifiably attained
legendary proportions among fossil aficionados in the western
United States. For purposes of propaedeutic pedagoguery, for
instance--instructing their students in the foundational principles
of Historical Geology--geology professors from community colleges
and universities (AKA, institutions of higher learning) throughout
the West regularly schedule field trips to the Vaughn Gulch exposures;
consequently, opportunities obviously arise there for lots of
folks to run off with loads of sample fossil material, although
one must note that at last field check it's still a productive
place to examine abundant showy, readily recognizable Paleozoic
Era sea life.
The Vaughn Gulch Limestone classically consists of about
1,500 feet of dominantly medium to dark gray limestone that additionally
incorporates subordinate shades of red, yellow, and orange; minor
shale partings typically separate the most diagnostic rock variety
present--a stunningly fossiliferous bioclastic limestone that
tends to form ledges crowded with the following forms: algae--Verticillopora
annulata; brachiopods--Gypidula, Athyris, Trematospira,
Schizophoria, Plectatrypa sp., Camarotoechia,
Atrypa, and Eatonia bicostata; such rugose and
tabulate corals as Australophyllum, Strombodes,
Favosites, Chonophyllitm, Rhisophyllum, Heliolites,
Alveolites, Cladopora, large cyathophyllids,
a small tryplasmid, a pycnostylid, Aulacophyllum,
Acinophyllum, Diplophyllum, Kyphophyllum nevadensis,
Phacelophyllum, Camerotoechia, Syringopora,
and Disphyllum; sponges--stromatoporoids and Hindia;
bryozoans; conodonts; and pelmatozoan and crinoidal echinoderms.
The youngest--and therefore final--significant fossil-bearing
unit within Mazourka Canyon's early to middle Paleozoice Era
successional complex is the lower to middle Devonian Sunday Canyon
Formation (around 415 million years old), which stratigraphically
speaking intertongues with and overlies the slightly younger
uppermost Vaughn Gulch sedimentary deposits. In its traditionally
familiar outcropping aspect, the Sunday Canyon weathers to form
rather poorly exposed slopes composed of thin flaggy fragments
of calcareous siltstone, calcareous shale, and argillaceous limestone
in shades of light gray to yellow and orange.
And it bears lots of invertebrate fossils. The Sunday Canyon
Formation faunal list, for example, includes at least six species
of the highly prized Monograptus graptolites, which developed
distinctive rhabdosomes that resemble tiny sawblades. This is
the westernmost outcropping of Mongraptus-producing strata
in the US, by the way. To find comparable, correlative lower
Devonian graptolite-yielding rocks to investigate, you'd have
to travel a few hundred miles east of Mazourka Canyon to the
Roberts Mountains Formation in central Nevada. Sunday Canyon
graptolite species present include: Monograptus dubius;
Monograptus tumescens; Monograptus vomerirnus;
Monograptus vulgaris; Monograptus scanius; Monograptus
uniformis, and Monograptus hercynious. Also well respresented
in the Sunday Canyon geologic sections are rynchonellid brachipods;
ostracods; sponge spicules; tentaculites--Tentaculites cf.
T. bellulus (taxonomic classification uncertain, but it could
be related to the modern-day pteropods, the sea snails); conodonts;
and corals--Alveolites, Favosites, Ceriod rugose
corals, cylindrical rugose corals, horn corals, Thamnopora
sp., and Cystiphyllum.
While fossil prospecting at Mazourka Canyon, it is fitting
to consider that before the great neighboring Sierra Nevada was
uplifted to its present impressive elevations, before it was
even a minor protuberance on the face of the earth, Mazourka
Canyon's fossil organisms now situated within its shadow had
already been covered over by countless primal ooze deposits at
the bottoms of unknown numbers of successive Paleozoic Era seas
some 541 to 252 million years ago. Eventually, through the ceaseless
invisible activity of passing vanished time, geologic forces
successfully thrust both the now lithified sedimentary beds of
those long-lived oceans and the younger, once-buried solidified
magma of the batholithic Sierra several thousands of feet above
sea level.
Now the inevitable, the inexorable laws of erosion take
their turn at the rocks. And yet, while standing atop Mazourka
Canyon's fossil beds--inspecting an outcrop of 485 to 415 million
year-old sedimentary rock, with the seemingly adamantine peaks
of the Sierra in bold relief against the western skyline--it
is perhaps difficult to believe that all mountain ranges, including
the surely eternal Sierra, must someday be no more. They must
be leveled to a plain, just as countless nameless ranges have
been so reduced in the geologic past.
But perhaps another sea will have its day atop their former
glory, and new creatures may stay alive in the rocks left behind,
to rise with the mountains of a yet-distant age to the delight
of future fossil hunters.
|